Prism: Spectral Self-Supervised Learning for Physiological Time Series

Clinical labels are often coarse, subjective, or arrive long after the signal that should have predicted them. A model trained to fit them inherits those limits, and at scale it learns to reproduce the labelling rather than the biology. This is why self-supervised learning suits physiological data: it learns directly from the signal, without a label needing to be attached.
Such a model still needs a term that prevents collapse; the trivial solution in which every input maps to the same point. SIGReg, an elegant recent construction from Balestriero and LeCunn, supplies one by drawing random directions in representation space and requiring that the embeddings projected onto each look like a standard Gaussian.
Physiological time series are not independent samples like images; they are trajectories in which the order carries the signal. SIGReg was built for independent samples, and so it does not see that order, which on this kind of data is precisely where the information lives.
Why time is different
SIGReg constrains the distribution of embeddings pooled across the batch and across time. That pooling across time is an issue. An image dataset is a collection of independent patches, so collecting them in a bag and asking what the bag looks like loses nothing. A physiological recording is not a bag of moments; it is a sequence, and the order is the signal. An ECG, a walking gait, a continuous blood-pressure waveform: the diagnostic content is the temporal structure, the rhythm and its irregularities, the cadence, the way one moment constrains the next. Pooling across the time axis tips that sequence into a bag and shakes it, and the regulariser only ever sees the bag.
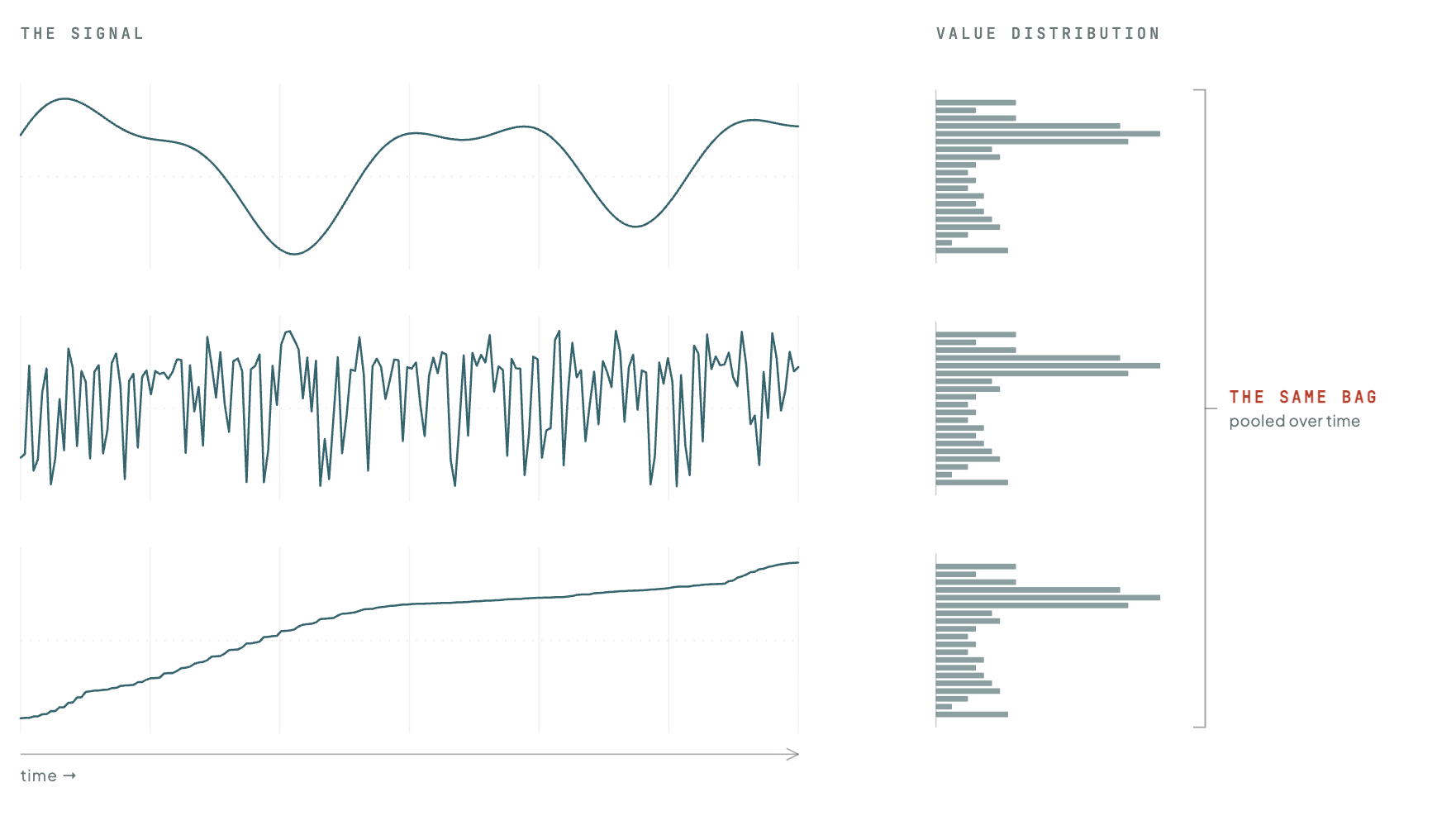
The cost of that is information. Anything that lives in how the signal changes over time, the rhythm, the cadence, the coupling between channels, leaves no trace in the bag, and a model under no pressure to keep it is free to let it go. The clearest case is the extreme one: an encoder can satisfy SIGReg perfectly and still discard the temporal structure entirely. Freeze each subject's recording to a single unchanging point, keep those points spread out across subjects, and the bag still looks Gaussian. Nothing in the objective objects, because the bag is all it checks.
This is not a failure the model would have to stumble into. A predictive model is trained to guess each next moment from the last, and the easiest signal to predict is one that never changes. So the training quietly pushes the encoder toward flat, frozen trajectories, and a term built for static inputs has no reason to resist, because the loss of structure does not show up in the bag.
Keeping time
SIGReg projects along random directions because space has no preferred frame, and one direction is as good as another. Time is different. It has a natural set of directions, its frequencies, and this is what Prism, the method we introduce here, is built on: it projects the time axis onto its Fourier basis rather than at random. Each window is decomposed into frequency components, and the same normality test is applied to those. Applied at a given frequency, the test reads the strength of the embedding's rhythm there, so the constraint now reaches the spectrum, the very structure the bag discarded. A frozen trajectory has no rhythm at all, no energy at any frequency, so where the pooled view cannot register the loss, the spectral test does, and the structure is preserved rather than allowed to drain away.
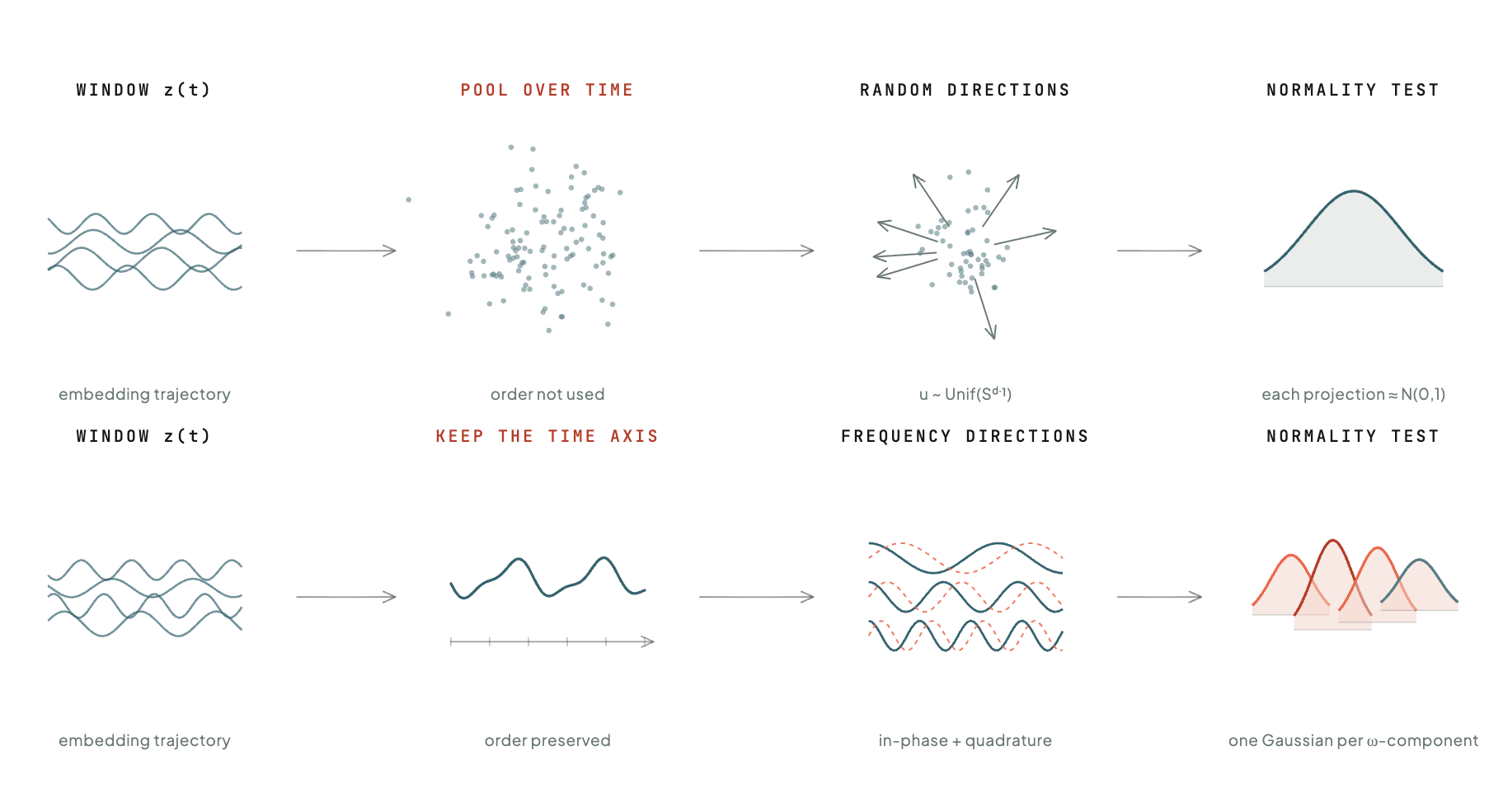
The target also has to be realisable: a regulariser of this kind is well-posed only if the distribution it asks for is one some process could actually produce. For a spectral target that condition is Herglotz's theorem, that a sequence is the autocovariance of a stationary signal exactly when its spectral density is nonnegative everywhere. It is the time-axis counterpart of the positivity condition the original method relied on in space. Prism rests on both at once, one spatial and one temporal, and it is the temporal one that certifies the construction.
From a theorem to physiology
The proof establishes that the gap exists and that the Fourier construction closes it. It says nothing about how much it matters in practice. In a controlled synthetic case the prediction is exact: Prism registers the loss of temporal structure by three orders of magnitude where plain SIGReg barely sees it.
We then tested it on four cohorts chosen to cover a range of physiological time series. Parkinsonian gait, where the signature is spectral and spread across an 18-channel force array. Atrial fibrillation, where it is a single-lead cardiac rhythm and the signal's amplitude alone carries almost nothing. Cardiovascular ageing, where heart-rate variability declines with age. And depression actigraphy as a control, where the signal is an overall activity level rather than a rhythm, so the correction should do nothing, which it does.
Prism improves discrimination consistently across the spectral cohorts and, as the theory demands, does no harm on the control. Two findings beneath that result are worth stating plainly. The advantage on gait runs through the overall spectrum rather than through coupling between channels: when we built a target that explicitly encoded that coupling, it did not reliably beat the simpler version, so we held the stronger claim back. And preserving a signal's spectrum preserves more of the subject's identity along with it, because each person's physiological rhythms are idiosyncratic, so Prism carries more subject and device identity than the plain version on the multichannel cohorts. For a representation meant to transfer across sites that is a real and useful thing to know, and it locates subject-invariance where it belongs, in a separate term rather than in the regulariser.
Our method
Prism is a good encapsulation of how we work. The target is upstream, complex but foundational: a regulariser suited to physiological time series, proven first in theory and then on real recordings, which improves our downstream models. Doing this well demands fluency in both the underlying mathematics and the ML. A more common path is to take a method built for images or text and apply it to physiological data; we start from the mathematics of the signal and adapt the method to it, here taking an elegant regulariser and extending it to also see time.
The same mathematical machinery carries across the continuous records of physiological state including cardiac rhythm, motion, respiration, cortical activity, glucose, each read through a different instrument and governed by related dynamics. Building measurements correctly, from the ground up, is the work we do at TimeTrace Labs.
The full paper is available on arXiv:
Please cite this work as:
TimeTrace Labs. Prism: Spectral Self-Supervised Learning for Physiological Time Series. TimeTrace Labs Blog, June 2026.
BibTeX:
@article{timetrace2026prism,
author = {{TimeTrace Labs}},
title = {Prism: Spectral Self-Supervised Learning for Physiological Time Series},
journal = {TimeTrace Labs Blog},
year = {2026},
url = {https://www.timetracelabs.com/blog/prism}
}